Тема 6. «Дыхание»
Задачи темы:
1) выяснить общее уравнение процесса дыхания и его значение в жизни
растительного организма;
1)
изучить историю
развития учения о дыхании;
2) изучить основные пути окисления дыхательного субстрата;
3) изучить этапы цикла гликолиза, ди - и трикарбоновых
кислот (цикл Кребса), глиоксилатного и
пентозофосфатного циклов, дыхательной ЭТЦ;
4) выяснить наличие взаимосвязи между дыханием и
брожением;
5) изучить хемиосмотическую
теорию сопряжения Митчелла;
6) изучить экологические и онтогенетические аспекты
дыхания;
Основные
понятия и термины темы: метаболизм,
катаболизм, анаболизм, гликолиз, цикл Кребса, глиоксилатный
и пентозофосфатный циклы, дыхательная электрон – транспортная цепь, эффект разобщения по Митчеллу,
пентозофосфатный шунт, дыхательный коэффициент.
ДЫХАНИЕ РАСТЕНИЙ
Живая клетка
представляет собой открытую энергетическую
систему,
она живет и сохраняет свою индивидуальность за счет постоянного притока
энергии. Как только этот приток прекращается, наступает дезорганизация и смерть
организма. Энергия солнечного света, запасенная при фотосинтезе в органическом
веществе, вновь высвобождается
и используется на самые различные процессы жизнедеятельности.
Энергетический цикл жизни можно представить в виде схемы (рис. 1).
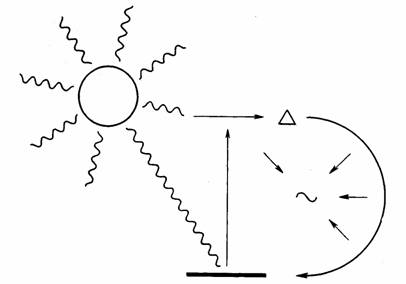
Рис.1. Энергетический цикл жизни
Как видно, энергия квантов света, аккумулированная в углеводах, вновь высвобождается в
процессе их распада (диссимиляции).
В самой общей форме можно отметить, что все живые клетки получают энергию за счет ферментативных реакций, в ходе
которых электроны переходят с более высокого энергетического уровня на более низкий. В природе
существуют два основных процесса, в
ходе которых энергия солнечного света, запасенная в органическом
веществе, высвобождается,— это дыхание и брожение. Дыхание
— это аэробный окислительный распад органических соединений на простые, неорганические, сопровождаемый выделением
энергии. Брожение —анаэробный процесс
распада органических соединений на более
простые, сопровождаемый выделением энергии.
При брожении степень окисленности соединений не
меняется. В случае дыхания акцептором электрона служит кислород, в
случае брожения — органические соединения. Процессы, входящие в энергетический цикл, имеют настолько важное значение, что в настоящее время возникла наука биоэнергетика, изучающая молекулярные и субмолекулярные основы
трансформации энергии.
ОБЩИЕ ВОПРОСЫ ДЫХАТЕЛЬНОГО ОБМЕНА
Суммарное уравнение
процесса дыхания:
С6H12O6 + 6O2 ->6СО2 + 6Н2О +
2824 кДж
Не вся
энергия, высвобождаемая при дыхании, может быть использована в процессах
жизнедеятельности. Используется организмом в основном та энергия, которая
аккумулируется в АТФ. Синтезу АТФ во многих случаях предшествует
образование" разности электрических зарядов на мембране, что, в свою
очередь, связано с разностью концентраций ионов водорода (протонный градиент или электрохимический градиент ионов Н+—∆μН +) по разные стороны мембраны. Согласно современным представлениям, не
только АТФ, но и ∆μН+ служат
источниками энергии для различных процессов жизнедеятельности
клетки (В. П. Скулачев). Обе формы энергии могут
быть использованы на процессы синтеза (химическая работа), процессы поступления, передвижения питательных
веществ и воды, процессы движения
(механическая работа), создание разности потенциалов между цитоплазмой и внешней средой (электрическая работа).
Энергия, не накопленная в ∆μН+ и
АТФ, в основном рассеивается в виде тепла, иногда света и для растения является
бесполезной. Выделение энергии в виде тепла
приводит к уменьшению упорядоченности
структур, т. е. к возрастанию энтропии.
1. ЗНАЧЕНИЕ ДЫХАНИЯ В ЖИЗНИ РАСТЕНИЯ
Дыхание — один из центральных
процессов обмена веществ растительного организма. Выделяющаяся при дыхании
энергия тратится как на процессы роста, так и
на поддержание в активном состоянии
уже закончивших рост органов растения. Вместе с тем значение дыхания не
ограничивается тем, что это процесс, поставляющий энергию. Дыхание,
подобно фотосинтезу, сложный окислительно-восстановительный
процесс, идущий через ряд этапов. На его промежуточных стадиях
образуются органические соединения, которые затем используются в различных
метаболических реакциях. К промежуточным
соединениям относят органические кислоты и пентозы, образующиеся при разных
путях дыхательного распада. Таким образом, процесс дыхания —
источник многих метаболитов. Несмотря на то,
что процесс дыхания в суммарном виде противоположен фотосинтезу, в некоторых случаях они могут дополнять
друг друга. Оба процесса являются
поставщиками как энергетических эквивалентов (АТФ, НАДФ.Н + ), так и метаболитов. Как видно из суммарного уравнения, в процессе дыхания образуется также
вода. Эта вода в крайних условиях
обезвоживания может быть использована растением и предохранить его от гибели. В некоторых
случаях, когда энергия дыхания
выделяется в виде тепла, дыхание ведет к бесполезной потере сухого вещества. В этой связи при рассмотрении процесса дыхания надо помнить, что не всегда усиление
процесса дыхания является полезным
для растительного организма.
2. АДЕНОЗИНТРИФОСФАТ. СТРУКТУРА И ФУНКЦИИ
Процессы обмена
вещества включают в себя реакции, идущие с потреблением энергии, и реакции с
выделением энергии. В некоторых случаях эти реакции сопряжены. Однако часто реакции,
в которых
энергия выделяется, отделены в пространстве и во времени от реакций, в которых
она потребляется. В процессе эволюции у растительных и животных организмов
выработалась возможность хранения энергии в форме соединений, обладающих богатыми
энергией
связями. Среди них центральное место занимает аденозинтрифосфат (АТФ). АТФ
представляет собой нуклеотидфосфат, состоящий из азотистого
основания (аденина), пентозы (рибозы) и трех молекул
фосфорной кислоты. Две концевые молекулы фосфорной кислоты образуют макроэргические,
богатые энергией связи. В клетке АТФ содержится главным образом в виде комплекса
с ионами
магния.
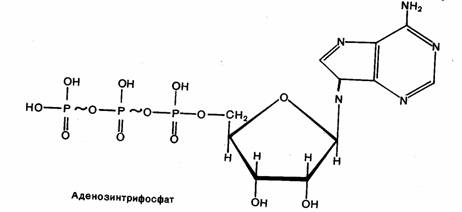
Рис.2. Структура АТФ
Аденозинтрифосфат в процессе дыхания
образуется из аденозиндифосфата и остатка
неорганической фосфорной кислоты (Фн) с
использованием энергии, освобождающейся при окислении различных органических веществ: АДФ + ФН→ АТФ + Н2О. При этом энергия окисления органических соединений
превращается в энергию фосфорной
связи.
В 1939—1940 гг. Ф. Липман установил, что АТФ служит главным переносчиком энергии в клетке. Особые свойства
этого вещества определяются тем, что
конечная фосфатная группа легко переносится с АТФ на другие соединения или отщепляется с выделением энергии, которая может быть использована на
физиологические функции. Эта энергия
представляет собой разность между свободной энергией АТФ и свободной энергией образующихся продуктов (∆G). ∆G —это изменение свободной энергии
системы или количество избыточной энергии,
которая освобождается при реорганизации
химических связей. Распад АТФ происходит по уравнению: АТФ + Н2О → АДФ + Фн, при этом происходит как бы
разрядка аккумулятора, при рН 7 выделяется ∆G = —30,6 кДж. Этот процесс катализируется ферментом аденозинтрифосфатазой
(АТФ-аза).
Равновесие гидролиза
АТФ смещено в сторону завершения реакции, что и обусловливает большую отрицательную
величину свободной энергии гидролиза. Это связано с тем, что при диссоциации четырех гидроксильных группировок при рН 7 АТФ имеет четыре отрицательных заряда (рис. 2).
Близкое расположение зарядов друг к другу
способствует их отталкиванию и, следовательно, отщеплению фосфатных
группировок. В результате гидролиза образуются соединения с одноименным
зарядом (АДФ3- и HPO42-),
которые отталкиваются друг от друга, что
препятствует их соединению. Уникальные свойства АТФ объясняются не только тем,
что при ее гидролизе выделяется большое количество энергии, но и тем, что она
обладает способностью отдавать концевую фосфатную группу вместе с запасом
энергии на другие органические соединения. Энергия, заключенная в макроэргической фосфорной связи,
используется на физиологическую
деятельность клетки. Вместе с тем по величине свободной энергии
гидролиза — 30,6 кДж/моль АТФ занимает промежуточное положение. Благодаря
этому система АТФ — АДФ может служить переносчиком
фосфатных групп от фосфорных соединений с более высокой энергией
гидролиза, например фосфоенолпируват (53,6 кДж/моль), к соединениям с более низкой энергией
гидролиза, например сахарофосфатам (13,8
кДж/моль). Таким образом, система АТФ — АДФ является как бы промежуточной
или сопрягающей.
3. ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНЫЕ ПРОЦЕССЫ.
РАБОТЫ А. Н. БАХА И В. И. ПАЛЛАДИНА
С химической точки зрения дыхание —
это медленное окисление. При
окислительно-восстановительных реакциях происходит перенос водорода или электрона от донора ДН2 (который
окисляется) к акцептору А (который
восстанавливается): ДН2+А→Д+АН2. Для того чтобы судить о направлении движения электронов между какими-либо двумя веществами, вводится понятие
стандартного восстановительного потенциала (Е0)
— это мера электронного давления. За нуль
потенциала условно принят восстановительный потенциал реакции Н2→2Н++2е-.
Чем более отрицательна величина восстанови- тельного потенциала, тем больше способность
данного вещества отдавать электроны (окисляться) или служить восстановителем. Наоборот, чем
положительнее величина восстановительного потенциала данного вещества, тем больше его способность
воспринимать электроны (восстанавливаться или служить окислителем). Восстановительный
потенциал кислорода равен +0,81В.
В
создании современных представлений о биологическом окислении большое значение имели работы двух крупнейших
русских ученых — В. И. Палладина (1859—1922) и А. Н. Баха
(1857—1946). Работы А. Н. Баха были посвящены возможности активации
кислорода воздуха. Молекулярный кислород — достаточно инертное соединение. Бах выдвинул предположение, что имеются ферменты-оксигеназы, активирующие кислород. Он считал, что процесс
активации состоит в том, что
происходит образование пероксидных соединений.
В. И. Палладии первый стал рассматривать дыхание как ряд ферментативных реакций. Основное значение в процессе
окисления он придавал процессу отнятия водорода от субстрата при участии
воды. Содержание своей теории В. И. Палладии выразил в виде следующих уравнений:
С6Н12О6 +
6Н2О + 12R → 6СО2 +12 RН2
12 RН2 + 602 → 12R +12Н20
С6Н12О6 +
6О2 -> 6СО2 + 6Н2О
Символом R Палладин обозначал дыхательный пигмент,
способный к обратимым
окислительно-восстановительным превращениям. Из приведенной схемы вытекают следующие важные положения: 1.
Непременным участником дыхания является вода. 2. Вода наряду с окисляемым субстратом выполняет роль
донора водорода. 3. В процессе дыхания участвуют специфические
активаторы водорода, отнимающие водород от субстрата. 4. Первые этапы дыхания являются анаэробными и не требуют присутствия
молекулярного кислорода. 5. Молекулярный кислород используется на заключительном этапе дыхания для регенерации акцепторов
водорода с образованием воды. Все указанные положения легли, как мы увидим, в основу современных представлений о процессе
дыхания, согласно которым дыхание идет в две фазы — анаэробную и аэробную, и молекулярный кислород используется на
регенерацию ферментов за счет Н+
воды и субстрата. В процессе дыхания активируется как водород субстрата, так и кислород воздуха.
4. СУБСТРАТЫ ДЫХАНИЯ
Вопрос о веществах,
используемых в процессе дыхания, издавна занимал физиологов. Еще в работах И. П.
Бородина было показано, что интенсивность процесса дыхания прямо пропорциональна содержанию в
тканях растений углеводов. Это дало основание предположить, что именно углеводы
являются основным веществом, потребляемым при дыхании. В выяснении данного
вопроса большое значение
имеет определение дыхательного коэффициента. Дыхательный коэффициент (ДК)
— это объемное или молярное отношение
СО2, выделившегося в процессе дыхания, к
поглощенному за этот же промежуток времени О2. При нормальном
доступе кислорода величина ДК зависит от
субстрата дыхания. Если в процессе дыханий используются углеводы, то
процесс идет согласно уравнению СбН12Сб + 6О2
= 6СО2 + 6Н2О. В этом случае ДК равен
единице.
Однако если разложению в процессе дыхания
подвергаются более окисленные соединения,
например органические кислоты, поглощение
кислорода уменьшается, ДК становится больше единицы. Так, если в качестве субстрата дыхания
используется яблочная кислота, то ДК = 1,33. При окислении в процессе
дыхания более восстановленных соединений,
таких, как жиры или белки, требуется больше кислорода и ДК становится меньше единицы. Так, при использовании
жиров ДК = 0,7. Определение дыхательных коэффициентов разных тканей растений показывает, что в нормальных условиях он близок к единице. Это дает основание считать,
что в первую очередь растение
использует в качестве дыхательного материала углеводы. При недостатке углеводов могут быть использованы и другие субстраты. Особенно это проявляется на проростках,
развивающихся из семян, в которых в
качестве запасного питательного вещества содержатся жиры или белки. В
этом случае дыхательный коэффициент
становится меньше единицы. При использовании в качестве дыхательного материала жиров происходит их расщепление до глицерина и жирных кислот. Жирные
кислоты могут быть превращены в углеводы через глиоксилатный
цикл. Использованию белков в качестве
субстрата дыхания предшествует их расщепление до аминокислот.
ПУТИ ДЫХАТЕЛЬНОГО ОБМЕНА
Существуют две
основные системы и два основных пути превращения дыхательного субстрата, или окисления углеводов: 1)
гликолиз + цикл Кребса (гликолитический); 2)
пентозофосфатный (апотомический). Относительная роль
этих путей дыхания может меняться в
зависимости от типа растений, возраста, фазы развития, а также в
зависимости от условий внешней среды. Процесс дыхания растений осуществляется во всех внешних условиях, в которых возможна
жизнь. Растительный организм не имеет приспособлений к регуляции
температуры, поэтому процесс дыхания осуществляется при температуре от
—50 до +50 °С. Нет приспособлений у растений и к поддержанию равномерного
распределения кислорода по всем тканям.
Именно необходимость осуществления процесса дыхания в разнообразных условиях привела к выработке в
процессе эволюции разнообразных путей дыхательного обмена и к еще
большему разнообразию ферментных систем,
осуществляющих отдельные этапы дыхания.
При этом важно отметить взаимосвязь всех процессов обмена в организме. Изменение пути дыхательного обмена
приводит к глубоким изменениям во всем
метаболизме растительных организмов.
1. ГЛИКОЛИТИЧЕСКИЙ ПУТЬ ДЫХАТЕЛЬНОГО ОБМЕНА
Данный путь
дыхательного обмена является наиболее распространенным и, в свою очередь, состоит из двух фаз. Первая
фаза — анаэробная (гликолиз), вторая фаза — аэробная. Эти фазы
локализованы в различных
компартментах клетки. Анаэробная фаза гликолиз
— в цитоплазме, аэробная фаза — в митохондриях.
Анаэробная фаза дыхания (гликолиз)
В процессе гликолиза
происходит преобразование молекулы гексозы до двух молекул пировиноградной кислоты: С6Н12О6->2СзН4О2
+ 2H2. Этот окислительный процесс может протекать в анаэробных условиях (в отсутствие кислорода) и идет
через ряд этапов. Прежде всего, для
того чтобы подвергнуться дыхательному распаду, глюкоза должна быть
активирована. Активация глюкозы происходит путем фосфорилирования шестого углеродного
атома за счет взаимодействия с АТФ.
Реакция идет в присутствии ионов магния и фермента
гексокиназы: глюкоза + АТФ→глюкозо-6-фосфат + АДФ. Затем глюкозо-6-фосфат изомеризуется
до фруктозо-6-фосфата. Процесс катализируется
ферментом фосфоглюкоизомеразой: глюкозо-6-фосфат→ фруктозо-6-фосфат. Далее происходит еще
одно фосфорилирование при участии АТФ. Фосфорная кислота присоединяется
к первому углеродному атому молекулы
фруктозы, процесс катализируется ферментом фосфофруктокиназой:
фруктозо-6-фосфат + АТФ→ фруктозо-1,6-дифосфат
+АДФ.
Дальнейшие реакции,
составляющие процесс гликолиза, складываются следующим образом:
фруктозо-1,6-дифосфат расщепляется с образованием двух триоз,
реакция катализируется ферментом альдолазой, которая
состоит из четырех субъединиц и содержит свободные SH-группы.
Реакция протекает по уравнению:
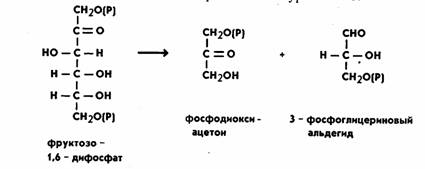
Молекула фосфодиоксиацетона при участии
фермента триозофосфатизомеразы превращается также в
3-фосфоглицериновый альдегид (ФГА). Дальнейшим превращениям подвергается именно
ФГА, окисляясь до 1,3-дифосфоглицериновой
кислоты (ДФГК). Это важнейший этап
гликолиза. Процесс идет с участием неорганического фосфата (H3РО4) и фермента глицеральдегид-3-фосфатдегидрогеназы.
Молекула этого фермента состоит из четырех
идентичных субъединиц. Каждая
субъединица представляет одиночную полипептидную цепь приблизительно из 220
аминокислотных остатков. Фермент содержит SH-группы
и кофермент НАД, который взаимосвязан с ферментом на всем протяжении процесса.
Сущность процесса заключается в окислении альдегидной группы ФГА в карбоксильную ДФГК. Окисление
идет с выделением энергии. За счет энергии окисления при участии неорганического фосфата (Н3РО4)
в молекуле ДФГК образуется
макроэргическая фосфатная связь. Одновременно происходит восстановление кофермента НАД.
В целом реакция выглядит следующим образом:
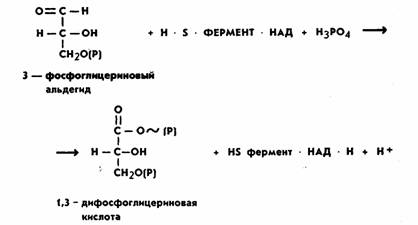
На следующем этапе за счет имеющейся макроэргической связи в 1,3-дифосфоглицериновой
кислоте образуется АТФ. Процесс катализируется
ферментом фосфоглицераткиназой:
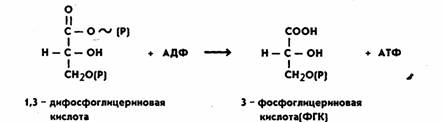
Таким образом, на этом этапе энергия окисления аккумулируется в форме
энергии фосфатной связи АТФ. Затем 3-ФГК превращается в 2-ФГК, иначе говоря, фосфатная группа
переносится из положения 3 в положение 2.
Реакция катализируется ферментом фосфоглицеромутазой и идет в присутствии магния:
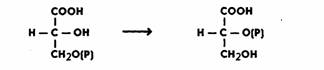
Далее происходит
дегидратация ФГК. Реакция идет при участии фермента енолазы в присутствии ионов Mg2+ или Мn2+. Дегидратация
сопровождается перераспределением энергии внутри молекулы, в результате чего возникает
макроэргическая связь. Образуется
фосфоенолпировиноградная кислота (ФЕП):
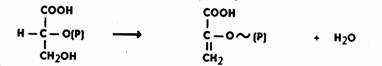
Затем фермент пируваткиназа переносит богатую
энергией фосфатную группу
на АДФ с образованием АТФ и пировиноградной кислоты.
Для протекания реакции необходимо присутствие ионов Mg2+ или Мn2+:
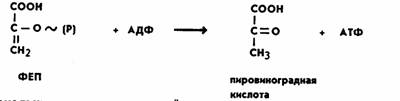
Поскольку при распаде одной молекулы
глюкозы образуются две молекулы
ФГА, то все реакции повторяются дважды. Таким образом, суммарное уравнение гликолиза следующее:
глюкоза + 2АТФ+ 2НАД+ + 2Фн + 4АДФ→2 пирувата + 4АТФ+ 2НАД.Н2 + 2АДФ.
В
результате процесса гликолиза образуются четыре молекулы АТФ, однако две из них
покрывают расход на первоначальное активирование субстрата. Следовательно,
накапливаются две молекулы АТФ. Образование АТФ в процессе
гликолиза носит название субстратного фосфорилирования,
поскольку макроэргические связи возникают на молекуле окисляемого субстрата.
Если считать, что при распаде
АТФ на АДФ и Фн выделяется 30,6 кДж, то за
период гликолиза накапливается в
макроэргических фосфатных связях всего 61,2 кДж. Прямые определения
показывают, что распад молекулы глюкозы до
пировиноградной кислоты сопровождается выделением 586,6 кДж.
Следовательно, энергетическая эффективность гликолиза невелика. Кроме того, образуется 2НАД.Н2. НАДН2 вступает в дыхательную цепь, что приводит к дополнительному образованию АТФ. Образовавшиеся две молекулы пировиноградной
кислоты вступают в аэробную фазу
дыхания.
Аэробная фаза дыхания
Вторая фаза дыхания — аэробная
— локализована в митохондриях и
требует присутствия кислорода. В аэробную фазу дыхания вступает пировиноградная кислота. Общее уравнение
этого процесса следующее:
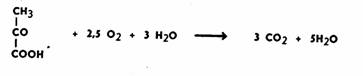
Процесс можно разделить на три основные
стадии: 1) окислительное декарбоксилирование
пировиноградной кислоты; 2) цикл трикарбоновых кислот (цикл Кребса); 3) заключительная
стадия окисления — электронно-транспортная цепь (ЭТЦ) требует
обязательного присутствия О2. Первые две
стадии происходят в матриксе митохондрий,
электронно-транспортная цепь локализована на внутренней мембране митохондрий.
Первая стадия — окислительное декарбоксилирование
пировиноградной кислоты. Общая формула
данного процесса следующая:
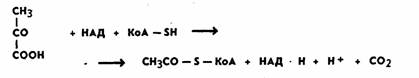
Процесс этот состоит
из ряда реакций и катализируется сложной мультиферментной
системой пируватдекарбоксилазой. Пируватдекарбоксилаза
включает в себя три фермента и пять коферментов (тиаминпирофосфат, липоевая кислота, коэнзим A (KoA-SH), ФАД и НАД. Вся эта система имеет молекулярную массу 4,0 . 106. В
результате этого процесса образуется активный ацетат — ацетилкоэнзим А (ацетил-КоА), восстановленный
НАД (НАД.Н + Н+), и выделяется углекислота (первая молекула).
Восстановленный НАД поступает в цепь переноса
электронов, а ацетил-КоА вступает в цикл трикарбоновых кислот. Важно отметить, что пируватдегидрогеназная система ингибируется АТФ. При
накоплении АТФ выше определенного
уровня превращение пировиноградной кислоты подавляется. Это один из способов регуляции интенсивности
протекания аэробной фазы.
Вторая стадия — цикл трикарбоновых кислот (цикл
Кребса). В
Общая схема цикла представлена на рисунке
3. В цикл вступает активный
ацетат, или ацетил-КоА. Сущность реакций, входящих в цикл, состоит в том, что ацетил- Ко А конденсируется с щавелевоуксусной кислотой (ЩУК). Далее превращение идет через ряд ди- и трикарбоновых
органических кислот. В результате ЩУК регенерирует в прежнем виде. В процессе
цикла присоединяются три молекулы Н2О,
выделяются две молекулы СО2 и четыре пары
водорода, которые восстанавливают
соответствующие коферменты (ФАД и НАД).
Суммарная
реакция цикла выражена уравнением: СН3СО — S —КоА + ЗН2О
+ ЗНАД + ФАД+АДФ + Фн →2СО2 + НS-КоА + ЗНАД.Н2 + ФАДН2 + АТФ. Отдельные реакции
протекают следующим образом. Ацетил-КоА, конденсируясь с ЩУК, дает лимонную кислоту, при этом КоА
выделяется в прежнем виде. Этот процесс катализируется ферментом цитратсинтетазой.
Лимонная кислота превращается в изолимонную. На следующем этапе происходит окисление
изолимонной кислоты, реакция катализируется ферментом
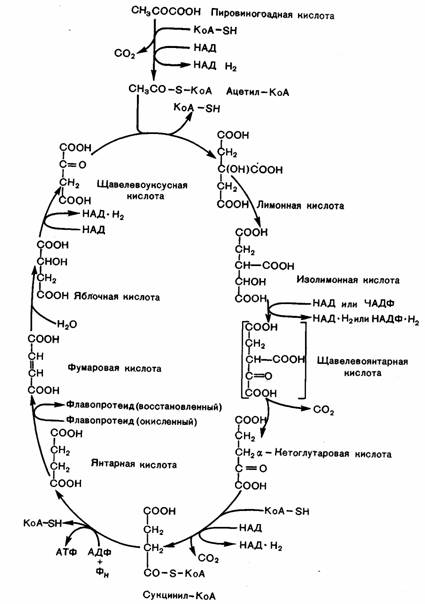
Рис. 3.
Цикл трикарбоновых кислот (цикл Кребса).
изоцитратдегидрогеназой. При этом водород
переносится на НАД (образуется НАД-РЬ). Для протекания этой
реакции требуются ионы магния или марганца. Одновременно происходит процесс декарбоксилирования. За счет одного из
атомов углерода, вступившего в цикл Кребса, первая молекула СО2 выделяется. Образовавшаяся кетоглутаровая
кислота подвергается окислительному декарбоксилированию подобно тому, которое
разбиралось по отношению к
пировиноградной кислоте. Этот процесс также катализируется мультиферментом
кетоглутаратдегидрогеназой, содержащей тиаминпирофосфат, липоевую кислоту, коэнзим А, ФАД и НАД. В результате
за счет второго атома углерода, вступившего в цикл, выделяется вторая молекула СО2.
Одновременно происходит восстановление еще
одной молекулы НАД до НАД-Ш и образуется сукцинил-КоА. На
следующем этапе сукцинил-КоА расщепляется на янтарную
кислоту (сукцинат) и HS — КоА.
Выделяющаяся при этом энергия накапливается в
макроэргической фосфатной связи АТФ. Этот этап важен, так как выделяющаяся энергия непосредственно накапливается в
АТФ. Этот тип образования АТФ, подобно ее образованию в процессе гликолиза,
относится к субстратному фосфорилированию. Образовавшаяся янтарная кислота окисляется до фумаровой кислоты.
Реакция катализируется ферментом сукцинатдегидрогеназой,
простетической группой которого является ФАД. Одновременно выделяется третья пара водородов, образуя ФАД-Н2.
На следующем этапе фумаровая
кислота, присоединяя молекулу воды,
превращается в яблочную кислоту. На последнем этапе цикла яблочная кислота окисляется до ЩУК. Эту
реакцию катализирует фермент малатдегидрогенеза,
активной группой которого является НАД, и
происходит выделение четвертой пары атомов водорода— образуется НАД-Ш. Таким образом, ЩУК регенерирует в прежнем виде и может реагировать со следующей
молекулой активного ацетата, поэтому практически ЩУК в процессе цикла не
расходуется. Одновременно в ходе каждого цикла выделяются две молекулы СО2 и образуются три молекулы НАД-Н + Н + и молекула ФАД-Н2. Многие реакции цикла
Кребса обратимы. Важно также
отметить, что образовавшиеся в рассмотренных реакциях органические
кислоты могут служить материалом для построения аминокислот, жиров и углеводов
и в этом случае выводятся из цикла. Вместе с тем соединения, входящие в цикл,
могут образовываться в ряде других реакций (например, при декарбоксилировании
аминокислот) и вступать в цикл. Таким
образом, рассмотренные превращения не
отделены от других реакций метаболизма, а тесно с ними взаимосвязаны. Для реакций цикла Кребса кислород
не требуется. Кислород необходим для регенерации или окисления
восстановленных коферментов (НАД-Н2
и ФАД-Ш). Количество окисленных форм этих коферментов ограничено. В
анаэробных условиях, когда регенерация
коферментов невозможна, они быстро оказываются исчерпанными и весь
процесс прекращается. Окисление коферментов осуществляется
в дыхательной цепи, или цепи переноса водорода и электронов. Конечным акцептором в этой цепи является кислород воздуха. Энергия, высвобождаемая при окислении
коферментов, накапливается в
макроэргических фосфатных связях АТФ.
Подводя
итог, можно отметить, что в результате распада 1 молекулы ПВК в аэробной фазе (декарбоксилирование ПВК + цикл Кребса) выделяется ЗСО2, 4НАД-Н2
и 1ФАД-Н2. Таким образом, 5 пар Н2,
образующихся из ПВК и воды, поступают в дыхательную
цепь.
Третья стадия —
электронно-транспортная цепь (ЭТЦ). В процессе окисления
пировиноградной кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем
рассматривать как 2Н + + 2е-. Именно в
таком виде они, акцептированные НАД и ФАД, и
передаются по цепи переносчиков. В процессе переноса водорода и электронов важную роль играют ферменты,
относящиеся к классу оксидоредуктаз.
Оксидоредуктазы, участвующие в
дыхательной цепи, делятся на следующие основные группы.
Пиридиновые дегидрогеназы, у которых коферментом служит НАД или НАДФ, отнимают
два атома водорода от субстрата. При этом к коферментам присоединяются один протон
и два электрона. Протон и один электрон связываются с атомом углерода в молекуле НАД, а второй электрон
нейтрализует положительный заряд атома азота. Один протон выделяется в среду (рис. 4). Коферменты НАД и НАДФ связаны с ферментом с помощью ионов металла и
сульфгидрильных группировок. В
зависимости от белкового носителя, к которому
присоединен кофермент (НАД или НАДФ), различают более 150 пиридиновых дегидрогеназ. Каждая из них специфична по отношению к определенному субстрату. Необходимо
учитывать, что НАД и НАДФ могут
воспринимать водород и электроны лишь в том случае, если субстрат имеет более отрицательное
значение потенциала по сравнению с
ними.
Флавиновые дегидрогеназы.
Это
также большая группа ферментов, катализирующая отнятие 2Н от различных
субстратов. Простетической группой этих ферментов
служат производные витамины В2 (рибофлавина) — флавинадениндинуклеотид
(ФАД) и флавинмононуклеотид (ФМН). Активной частью флавиновых
дегидрогеназ служит изоаллоксазиновое
кольцо. В процессе восстановления
именно к этой группировке присоединяется 2Н (2Н++2е) (рис.5).
Простетическая группа у флавиновых
дегидрогеназ прочно прикреплена
к белковому носителю. Специфичность и в этом случае определяется
белковой частью фермента.
Цитохромы. Простетическая группа цитохромов представлена железопорфиринами. Железопорфириновая
группа (тем) в цитохромах прочно связана с
белком через атомы серы аминокислоты цистеина. Известно около 20 цитохромов,
которые делятся на четыре главных класса: а, Ь, с, d, отличающихся между собой природой простетической группы: цитохромы
а содержат железоформилпорфирины, цитохромы b — железопротопорфирины,
цитохромы d — железогидропорфирины. В каждую группу цитохромов
входит по нескольку различающихся
между собой ферментов.
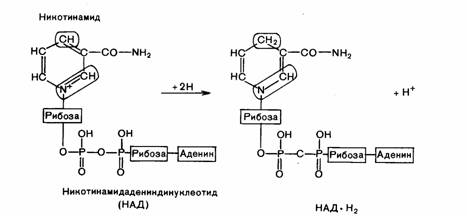
Рис. 4. Окисленная и восстановленная форма никотинамидадениндинуклеотида (НАД)
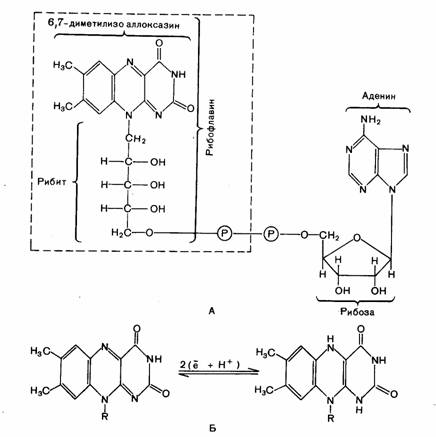
Рис.5. А - Структурные формулы флавиновых
нуклеотидов: ФМН ( в рамке) и ФАД (вся формула); Б -
восстановление и дегидрирование
флавиновых нуклеотидов
Роль цитохромов заключается в переносе электронов. Содержащееся
в цитохромах железо способно к обратимым
окислительно-восстановительным реакциям. Воспринимая электрон, железо
восстанавливается, теряя его
— окисляется: Fe3+ +е-→ Fe2+.
Непосредственно с кислородом воздуха может
реагировать только цитохромоксидаза, которая кроме железа содержит атомы меди.
Помимо
перечисленных ферментов в переносе электронов по дыхательной цепи принимают участие кофермент
Q и железосерные белки.
Кофермент Q
—это производное бензохинона, получившее название убихинон. Убихинон представляет собой кольцевую молекулу с двумя
присоединенными к ней атомами кислорода, для которой возможны три состояния. В
полностью окисленном состоянии, или хиноновой форме, оба атома кислорода связаны с кольцом двойными связями.
Присоединение одного атома водорода к одному из атомов кислорода дает полухиноновую форму QH. В полностью восстановленной форме атомы водорода присоединяются к обоим
атомам кислорода. Эта форма носит название гидрохиноновой
— QH2 (рис. 6).
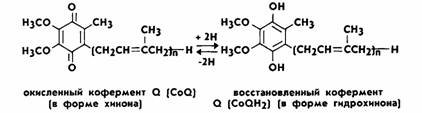
Рис.6. Окисленная и восстановленная формы коферментов
Q
Таким образом, кофермент Q может присоединять 2 протона и 2
электрона. Убихинон растворим в жирах и в связи с
этим подвижен в липидной фазе мембран. Железосерные белки
содержат FeS — это переносчики электронов
подобно цитохромам. Содержащееся в них железо обратимо восстанавливается и окисляется.
Путь
переноса водорода и электрона от одной молекулы переносчика к другой (рис. 7) представляет
собой окислительно-восстановительный
процесс. При этом молекула, отдающая электрон или водород, окисляется, а молекула, воспринимающая
электрон или водород,
восстанавливается. Движущей силой транспорта водорода в дыхательной цепи является разность потенциалов. В
связи с этим расположение отдельных переносчиков в дыхательной цепи, так же как
и в цепи фотосинтетической,
определяется величиной их окислительно-восстановительного потенциала (О/В). В начале цепи расположен НАД, обладающий наибольшей отрицательной величиной О/В потенциала (—0,32 В), а в конце —
кислород с наиболее положительной величиной (+0,82 В). Остальные переносчики
ФАД, KoQ цитохромы расположены между
ними в порядке последовательного повышения положительного потенциала. Это и
позволяет электронам передвигаться по направлению к кислороду (наивысший положительный потенциал).
В 1939—1940 гг. биохимик В. А. Белицер указал, что выделяющаяся в процессе передачи по дыхательной цепи электронов энергия
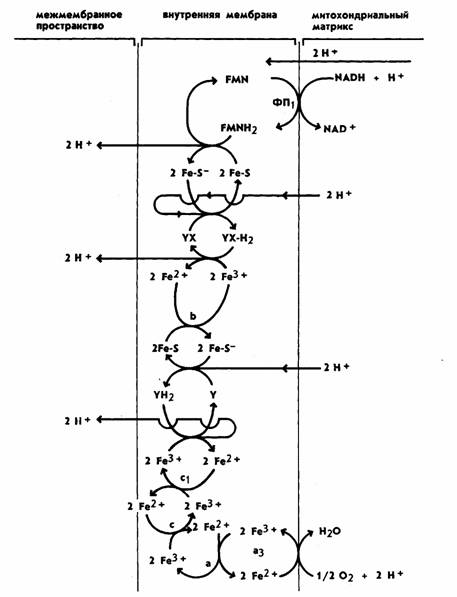
Рис. 7. Дыхательная цепь (расположение переносчиков в
мембране митохондрий)
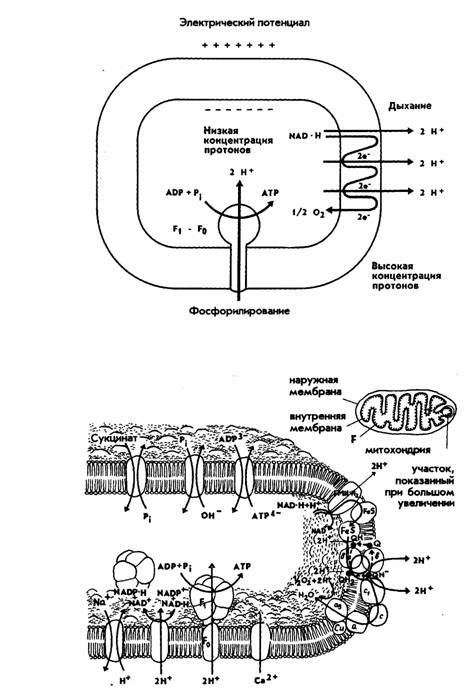
Рис. 8. Окислительное фосфорилирование
в митохондриях: А – схема Митчелла; Б – расположение переносчиков в
мембране митохондрий
частично
накапливается в АТФ. При переносе электронов свободная энергия системы
постепенно уменьшается. Общее изменение энергии при переносе пары водородов и
пары электронов от НАД на кислород можно рассчитывать по формуле: ∆Go = nF∆Eo, где п — число электронов,
равное 2; F —
фарада = 96 633,97 Дж; ∆ E0 — разность потенциалов между участком цепи от —0,32
до +0,82= 1,14; ∆ Go — стандартное изменение свободной энергии, ∆ Gо = 2-96633,97 Дж-1,14 = 220,8 кДж.
Таким образом,
изменение свободной энергии системы составляет около 220,8 кДж. Свободная энергия гидролиза АТФ равна 30,6
кДж. Исходя из того, что уменьшение свободной энергии системы при переносе пары электронов с НАД на
кислород составляет 220,8 кДж, можно было предположить возможность
образования из АДФ + Фн семи молекул АТФ.
Однако было показано, что при прохождении пары электронов от НАД-Н до 1/2 О2 образуется всего 3 молекулы АТФ. Из этого был
сделан вывод, что в цепи переноса электронов
имеется три пункта фосфорилирования. Накопление
энергии окисления в АТФ при продвижении электрона по цепи переносчиков называют окислительным фосфорилированием.
Механизм образования АТФ в процессе окислительного фосфорилирования, так же как и фотофосфорилирования,
объяснен благодаря работам
английского биохимика П. Митчелла. Его теория получила название хемиосмотической.
Согласно хемиосмотической теории свободная энергия, образованная при
окислительно-восстановительных реакциях в дыхательной цепи, преобразуется в
электрохимический градиент ионов водорода (∆μН
+). При этом мембрана переходит в высокоэнергетическое
состояние. Ионы Н+ (протоны) переносятся с внутренней стороны внутренней мембраны на ее
внешнюю сторону, иначе говоря, из
матрикса митохондрии в межмембранное пространство с помощью
переносчиков. ∆μН +, в свою
очередь, является источником энергии для
образования АТФ из АДФ. ∆μН+ имеет две составляющие: градиент рН (∆ рН) и градиент электрического потенциала. Переносчики дыхательной
цепи сосредоточены на внутренней мембране
митохондрий. При этом они как бы вплетены в митохондриальную мембрану и составляют дыхательные ансамбли. Так же
как в мембранах хлоропластов,
переносчики, расположенные в митохондриях,
неоднородны. Одни из них переносят целые атомы водорода, а другие
— лишь электроны. Использование переносчиков второго типа (переносящих электрон) возможно потому, что протоны (Н+) могут находиться в водной среде
клетки в свободном состоянии. В
мембране митохондрий, так же как и в мембране хлоропластов, переносчики
водорода чередуются с переносчиками электрона, что имеет принципиальное значение для хемиосмотической теории. Молекула переносчика, несущая целый атом водорода, взаимодействует с переносчиком, воспринимающим только электроны, и
протоны высвобождаются в межмембранное
пространство. Именно это, согласно хемиосмотической теории, лежит в основе преобразования выделяющейся в процессе окисления энергии в
энергию электрохи мического мембранного потенциала
— ∆μН+ и далее в энергию
АТФ. Согласно теории Митчелла, при переносе
пары электронов от НАД на кислород они пересекают мембрану 3 раза и этот перенос сопровождается выделением на внешнюю
сторону мембраны 6 (3 пар) протонов (рис. 8, А, Б).
При рассмотрении рисунков 7 и 8 надо
учитывать, что это лишь предположительная
схема расположения переносчиков. Многое в ней нуждается в уточнении. Как видно
из приведенной схемы, восстановленный
кофермент НАДН+ +Н+, образующийся в реакциях цикла Кребса, располагается на внутренней стороне мембраны
митохондрий. На первом этапе ФАД
воспринимает водороды от НАД и восстанавливается,
образуя ФАД-Н2. С помощью этого фермента 2Н + переносятся на
другую (внешнюю) сторону мембраны, и здесь происходит первое
разделение зарядов. Два протона (2 Н+) выделяются на внешнюю сторону внутренней мембраны, а электроны
присоединяются к переносчику
(железосерный белок), с помощью которого переносятся на внутреннюю сторону мембраны. При этом происходит восстановление
железа (Fe3++e-→Fe2+). Этот
переносчик переправляет электроны снова на
внутреннюю сторону мембраны. Здесь электроны акцептируются KoQ (убихинон — переносчик Н), который, заряжаясь отрицательно, захватывает двумя
электронами два протона из внутренней среды. Поскольку KoQ растворим в
липидах, он диффундирует к внешней стороне мембраны и выделяет там еще 2Н + (второе разделение зарядов), а электроны
передаются на цитохром b. Рассматривая схему, мы указали на два места выделения (всего четырех) протонов.
Между тем, согласно хемиосмотической теории, локализация
пунктов фосфорилирования в дыхательной цепи определяется пунктами выделения ионов Н +. Поскольку, как указывалось выше, показано наличие трех мест фосфорилирования, то необходима транслокация через внутреннюю
мембрану трех пар протонов. Однако
точно место выделения третьей пары протонов не установлено. Предполагается, что третья пара Н+
выделяется также при переносе электронов от KoQ (убихинона) к цитохрому b. При
этом участвуют 2 молекулы убихинона, которые сначала
переходят в полухинон, а затем в гидрохинон (выделяется третья пара Н+).
Далее электроны
передвигаются по цепи цитохромов b, с, а,
а3, содержащих железо. В каждом из них происходят обратимые окислительно-восстановительные превращения железа. На заключительном этапе электроны переносятся ферментом цитохромоксидазой (содержащей наряду с железом медь) на внутреннюю сторону
мембраны на кислород. Кислород,
заряжаясь, воспринимает 2Н+ из внутренней среды с образованием Н2О. В результате выброса ионов Н+
на внешнюю сторону мембраны
митохондрий и создается электрохимический градиент протонов (∆μН+). Таким образом, сам механизм
процессов, происходящих на мембранах
хлоропластов и митохондрий, сходен. Однако имеются два основных отличия:
1) в случае хлоропластов источником энергии
потока электронов служит энергия света, а у митохондрий — энергия
окислительных процессов; 2) распределение протонов
на мембране противоположно: у митохондрий протоны - накапливаются на наружной стороне, а у
хлоропластов — на внутренней.
Протонный градиент
представляет собой как бы резервуар свободной энергии. Эту энергию можно использовать
при обратном потоке протонов через мембрану. При этом происходит разрядка мембраны. В
частности, энергия может быть затрачена на синтез АТФ. Процесс синтеза
АТФ идет с помощью специальных ферментов — АТФ-синтетаз (АТФ-азы). Этот фермент обратимого действия в зависимости от
условий может осуществлять не только синтез АТФ с поглощением, но и ее гидролиз с выделением энергии. АТФ-азы локализованы
на мембранах в виде грибовидных частиц, состоящих из «шляпки», выступающей на поверхности
мембраны, и «ножки», погруженной в толщу мембраны. Активный участок
фермента расположен в
«шляпке», а «ножка» представляет собой канал, по которому
осуществляется обратный — по градиенту концентрации ток протонов.
Существуют
две гипотезы, объясняющие механизм синтеза АТФ прямой и косвенный. Согласно прямому механизму, АДФ и Фн связываются с активным центром фермента, куда по
каналу поступают протоны. Два протона
соединяются с кислородом фосфата с образованием Н2О. Это
делает Фн активным, и он присоединяется к АДФ. После этого молекула АТФ отделяется от
фермента. Согласно второй гипотезе, АДФ присоединяет молекулу Фн в активном центре фермента спонтанно.
Однако образующаяся при этом молекула АТФ прочно
связывается с ферментом, и для ее освобождения требуется энергия.
Энергия доставляется протонами, которые, воздействуя на фермент, изменяют его конформацию, благодаря
чему АТФ высвобождается.
Доказательством
того, что именно градиент протона обеспечивает фосфорилирование, являются опыты с разобщителями
окисления и фосфорилирования. Как уже упоминалось, к таким разобщителям
относится динитрофенол. Оказалось, что
действие динитрофенола связано с тем, что он делает мембрану проницаемой для протонов и тем самым ликвидирует протонный градиент. При
этом скорость окисления даже усиливается, однако фосфорилирование
не происходит. Таким образом, процесс окисления сопряжен с процессом фосфорилирования. Степень сопряженности окисления и фосфорилирования может быть разной в зависимости от условий
и от состояния клеток. Показателем сопряженности окисления и фосфорилирования служит коэффициент фосфорилирования
Р/О. Это — отношение количества связанного
неорганического фосфора (АДФ + Фн →АТФ) к поглощенному в процессе дыхания
кислороду. Как уже рассматривалось выше, перенос двух электронов к кислороду по
дыхательной цепи сопровождается не более чем тремя фосфорилированиями.
Следовательно, коэффициент Р/О может быть не более 3.
На величину Р/О
оказывают влияние внешние условия. При засухе окисление усиливается, а накопление энергии в виде АТФ не
происходит, коэффициент Р/О резко падает. Коэффициент фосфорилирования
резко падает и при заболевании
организмов.
В ряде случаев может
наблюдаться непосредственное использование энергии протонного градиента (∆μН+).
Действительно, поскольку внутренняя сторона
мембраны оказывается заряженной отрицательно,
возникает трансмембранный потенциал. Катионы в силу электрического притяжения могут поступать и
накапливаться во внутреннем
пространстве митохондрий. Имеются данные, что протонный градиент может обеспечить также приток
углеводов, в частности поступление
сахарозы в ситовидные трубки. Таким образом, ∆μН+
обеспечивает осмотическую работу и транспорт веществ против градиента их концентрации. Наконец, показана
возможность использования ∆μН+
на механическую работу (движение бактерий). Вместе
с тем важно отметить, что ∆μН+
может играть роль как транспортная форма энергии, передаваясь вдоль
мембран (В. П. Скулачев).
Таким образом,
клетка обладает двумя формами используемой энергии, двумя энергетическими «валютами» — АТФ
и ∆μН+:1) АТФ —
химическая «валюта», растворимая в воде и легко используемая в
водной фазе; 2) ∆μН+ —
электрохимическая, неразрывно связанная с
мембранами. Важно заметить, что эти две формы используемой клеткой энергии могут переходить друг в друга. При образовании
АТФ используется энергия ∆μН+
при распаде АТФ энергия может аккумулироваться в∆μН+:
Энергетический баланс процесса дыхания
Подводя итоги
энергетики процесса дыхания, подсчитаем, сколько всего молекул АТФ может
образоваться при распаде одной молекулы глюкозы. В первую анаэробную фазу дыхания
при распаде одной молекулы глюкозы до двух молекул пировиноградной кислоты в процессе
субстратного фосфорилирования накапливаются две молекулы АТФ.
Одновременно на этой фазе дыхания при окислении ФГА до ФГК в цитозоле
образуются две молекулы восстановленных
коферментов (2 НАД-Н2). Они диффундируют через наружную мембрану и окисляются в дыхательной цепи благодаря наличию у растений НАД-Н-дегидрогеназы,
локализованной на наружной поверхности
внутренней мембраны. При этом синтезируется 6 молекул АТФ. В аэробной
фазе дыхания при окислении пировиноградной
кислоты образуются 4НАД-Н2. Их окисление в дыхательной цепи приводит к
образованию 12 АТФ. Кроме того, в цикле
Кребса восстанавливается одна молекула флавиновой дегидрогеназы (ФАД-Н2). Окисление этого соединения в дыхательной
цепи приводит к образованию 2 АТФ, поскольку одно фосфорилирование не происходит. При окислении молекулы а-кетоглутаровой кислоты до янтарной энергия непосредственно
накапливается в одной молекуле АТФ (субстратное фосфорилирование).
Таким образом, окисление одной молекулы
пировиноградной кислоты
сопровождается образованием ЗСО2 15 молекул
АТФ. Однако при распаде молекулы глюкозы
образовались две молекулы пировиноградной
кислоты. Следовательно, всего в аэробной фазе дыхания образуется 6
молекул СО2 и 30 молекул АТФ плюс 8АТФ в
анаэробной фазе. Итого 6 молекул СО2 и 38
молекул АТФ образуется в процессе
окислительно-дыхательного распада молекулы гексозы. На образование 38
молекул АТФ затрачено 38-30,6 кДж = 1162,8 кДж. Всего при сжигании 1 моль
глюкозы выделяется 2824 кДж: C6H12O6 + 6O2->6CO2 + 6H2O + 2824 кДж. Таким образом, КПД процесса дыхания при самых благоприятных
условиях составляет около 40%.
Подводя итоги, можно
сказать, что биологическое окисление — это
многоступенчатый ферментативный процесс, сопровождаемый выделением энергии.
Химизм процесса брожения
Брожение — это внутренний
окислительно-восстановительный процесс, при котором акцептором электронов
служит органическая молекула и суммарная степень окисления образующихся
продуктов не отличается от степени
окисления сбраживаемого вещества. С. П. Костычев выдвинул
положение о генетической связи процессов брожения и дыхания. При этом он опирался на
следующие факты:
1.У высших растений был найден весь набор
ферментов, который катализирует отдельные этапы процесса брожения.
2. При временном
попадании в условия анаэробиоза высшие растения определенное время существуют за
счет энергии, выделяющейся в процессе брожения. Правда, поскольку процесс брожения энергетически
значительно менее эффективен, в анаэробных условиях рост растений
приостанавливается. Кроме того, продукты брожения, в частности спирт, ядовиты, и их
накопление приводит к гибели растения.
3. При добавлении к
клеткам факультативных анаэробов (дрожжи) полусброженных
Сахаров интенсивность дыхания у них резко возрастает, следовательно, полусброженные продукты являются лучшим субстратом
дыхания по сравнению с неизмененными сахарами.
В настоящее время
общепризнано, что первые этапы (гликолиз) протекают одинаково при процессах как дыхания, так и брожения. Поворотным моментом является образование пировиноградной кислоты. В аэробных условиях
пировиноградная кислота распадается
до СО2 и воды в результате декарбоксилирования и цикла Кребса (дыхание), тогда как в анаэробных она преобразуется в
различные органические соединения (брожение). Организм обладает способностью при изменении условий переключать процессы, прекращая
брожение и усиливая дыхание и наоборот. Впервые в опытах Пастера было показано, что в присутствии кислорода процесс
брожения у дрожжей тормозится и заменяется процессом дыхания. Одновременно резко сокращается распад глюкозы. Это явление
оказалось характерным для всех факультативных анаэробных организмов, включая и высшие растения, и
получило название эффекта Пастера.
Сокращение расхода глюкозы в присутствии кислорода целесообразно, поскольку при дыхательном распаде выход энергии
значительно выше, а следовательно, глюкоза используется более экономно. Однако осуществление
разбираемого эффекта требует
специальных механизмов, которые будут рассмотрены далее.
В
зависимости от получаемого продукта различают разные типы брожения. При спиртовом брожении пировиноградная
кислота, образовавшаяся в процессе гликолиза,
декарбоксилируется с образованием уксусного
альдегида при участии фермента пируватдекарбоксилазы, а затем восстанавливается до этилового спирта ферментом алкогольдегидрогеназой:
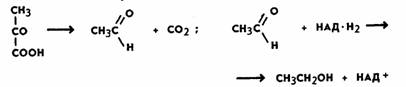
Ни та
ни другая реакция не сопровождается образованием АТФ. В связи с этим выход
АТФ при спиртовом брожении такой же, как при гликолизе (первой фазы брожения и
дыхания), и составляет две молекулы при распаде 1 моль глюкозы. Восстановленные
никотинамидные
коферменты (НАД-Н2), образовавшиеся в процессе гликолиза, не
поступают в дыхательную цепь (у анаэробных организмов ее и нет), а используются для
восстановления уксусного альдегида до спирта. Следовательно, энергетический выход
процессов брожения крайне
низок. Разные микроорганизмы осуществляют и разные
типы брожения. Так, молочнокислые бактерии накапливают молочную кислоту. При этом пировиноградная кислота
восстанавливается до молочной
кислоты:
|
|
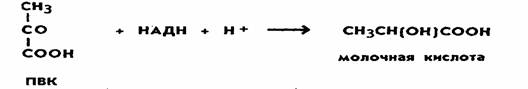
Для некоторых оолигатных анаэробных микроорганизмов, например азотфиксирующей бактерии Clostridium pasteurianum, характерно
образование в процессе брожения масляной кислоты (СН3СН2СН2СООН),
СО2 и Н2. Процесс брожения
служит источником энергии для облигатных
(обязательных) или факультативных анаэробных
организмов.
2. ПЕНТОЗОФОСФАТНЫИ ПУТЬ ДЫХАТЕЛЬНОГО ОБМЕНА
Наряду с разложением
глюкозы в процессе гликолиза существует другой, не менее распространенный путь —
пентозофосфатный, или апотомический. В этом случае
глюкоза также подвергается первоначальному фосфорилированию с образованием глюкозо-6-фосфата,
затем пути расходятся. Монофосфорный эфир глюкозы подвергается окислению при участии фермента глюкозо-б-фосфат- дегидрогеназы.
Коферментом является НАДФ, при этом образуется фосфоглюконовая кислота. Процесс происходит следующим образом:
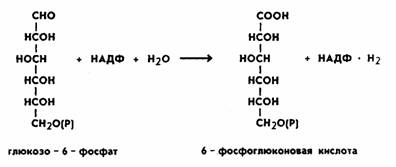
На следующем этапе происходит процесс
окислительного декарбоксилирования фосфоглюконовой кислоты. В результате
этой peaкции отщепляется одна
молекула углекислого газа и образуется пятиуглеродный
сахар рибулезо-5-фосфат. Отсюда и название пентозофосфатный цикл или апотомический
путь (апотомия — усекновение). Реакция катализируется ферментом фосфоглюконатдегидрогеназой с коферментом НАДФ:
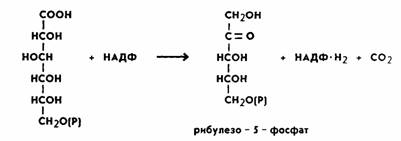
Обе рассмотренные реакции необратимы.
Последующие реакции представляют цикл. Для прохождения полного цикла реакций необходимы три молекулы гексозомонофосфата, которые превращаются в три молекулы
рибулезо-5-фосфата по уравнению: 3-гексозомонофосфата + 6 НАДФ +ЗН2О →3 пентозофосфата
+6 НАДФ-H2 + ЗСО2. Три образовавшиеся молекулы пентозы
(рибулезо-5-фосфата) претерпевают превращения.
Первая молекула рибулезо-5-фосфата под
влиянием фермента эпимеразы превращается в
ксилулозо-5 фосфат. Вторая молекула
рибулезо-5-фосфата превращается в рибозо-5-фосфат
под влиянием фермента фосфопентоизомеразы. Ксилулозо-5-фосфат реагирует с рибозо-5-фосфатом, в
результате получается семиуглеродный сахар
— седогептулозо-7-фосфат и фосфоглицериновый
альдегид. Эта последняя реакция идет под влиянием фермента транскетолазы:
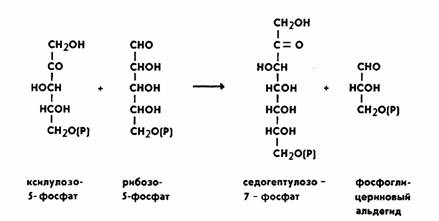
Семиуглеродный сахар реагирует с
ФГА, при этом образуется фруктозо-6-фосфат и эритрозофосфат.
Фруктозо-6-фосфат изомеризуется до глюкозо-6-фосфата.
Эритрозофосфат вместе с третьей молекулой рибулезо-5-фосфата также дает
глюкозо-6-фосфат и ФГА. В целом процесс представлен на рисунке 68, а также
может быть суммирован в следующем виде:
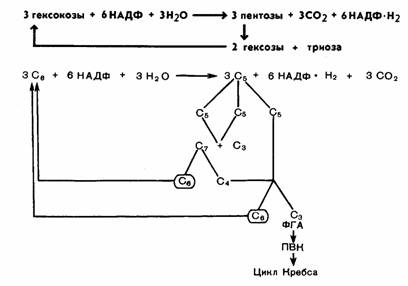
Триоза (ФГА) может
претерпевать превращения по гликолитическому пути до пировиноградной кислоты и
далее вступать в цикл Кребса. Как видно из уравнения, в пентозофосфатном цикле
из каждых
трех молекул глюкозы распадается одна, при этом выделяются 3 молекулы СО2
и образуются 6 НАДФ-Н2 и ФГА. Окисление НАДФ-Н2 идет через дыхательную цепь, при этом первоначально
НАДФ-Н2 взаимодействует с НАД. При
включении 6 НАДФ-Н2 в дыхательную цепь образуется 18 молекул АТФ.
Если считать, что 1 молекула АТФ используется
на предварительное фосфорилирование гексозы, то накапливается 17 молекул АТФ.
Вместе с тем при превращении
фосфоглицеринового альдегида в пировиноградную кислоту и ее окислении через цикл Кребса образуется еще 20
молекул АТФ. Таким образом, при окислении по пентозофосфатному пути одной
молекулы глюкозы выделяется 37 молекул АТФ. Следовательно,
с энергетической стороны апотомический путь почти не отличается от гликолитического.
Однако они сильно различаются по
промежуточным продуктам. В процессе апотомического распада глюкозы образуется НАДФ-Н2,
который может быть использован на
различные синтетические процессы, в том числе и в темновых реакциях фотосинтеза. Важное значение имеет
образование пентоз. Пентозы входят в
состав нуклеиновых кислот. Нельзя не
отметить, что образующийся в этом процессе эритрозофосфт
является предшественником лигнина и
ряда других соединений Внимательное
рассмотрение реакций окисления глюкозы, связанных с пентозофосфатным путем, показывает его большое сходство с превращением углеводов в фотосинтетическом цикле Кальвина Возможно,
что благодаря наличию одинаковых промежуточных продуктов эти процессы в ряде пунктов взаимосвязаны.
В этой связи интересно отметить, что
наряду с локализацией пентозофосфатного пути в цитоплазме он может в отсутствие света осуществляться и в хлоропластах. На протяжении онтогенеза растений
и в зависимости от условий происходит смена дыхательных путей. В силу образования различных промежуточных продуктов это
оказывает значительное влияние на обмен веществ и физиологическую деятельность растительного организма. Существует ряд
методов, с помощью которых можно установить соотношение путей дыхательного обмена (применение специфических ингибиторов,
метод с использованием меченых атомов
и др.). Однако все они имеют свои недостатки, и в силу этого к
экспериментальным данным по этому вопросу
надо подходить с большой осторожностью.
ВЛИЯНИЕ ВНЕШНИХ И ВНУТРЕННИХ ФАКТОРОВ НА ИНТЕНСИВНОСТЬ ДЫХАНИЯ
Показатели
интенсивности дыхания прямо противоположны показателям интенсивности фотосинтеза.
Интенсивность дыхания можно
определить: 1) по количеству выделенного СО2;
2) по количеству поглощенного кислорода; 3) по убыли сухой массы. Все эти три показателя
рассчитываются на единицу массы в единицу времени
1. ВЛИЯНИЕ ВНЕШНИХ УСЛОВИЙ НА ПРОЦЕСС ДЫХАНИЯ
Температура. Дыхание у ряда
растений осуществляется и при температуре ниже О °С. Так, у
хвои ели процесс дыхания идет даже при температуре — 25 °С. Как всякая ферментативная реакции с
повышением температуры интенсивность дыхания возрастает. Однако это происходит до определенного предела,
выше которого начинается инактивация ферментов и интенсивность дыхания снижается. При этом надо учитывать длительность
выдерживания растения при данной
температуре. При кратковременной экспозиции интенсивность дыхания
возрастает при повышении температуры до 35 °С и даже
40 °С. При длительном выдерживании в такой температуре
интенсивность дыхания уменьшается. Для суждения о влиянии температуры на какой-либо процесс обычно
оперируют величиной температурного
коэффициента. Температурный коэффициент (Q10) процесса дыхания зависит от типа растений и от градаций температуры.
Так, при повышении температуры от 5 до 15°С Q10 может возрастать до 3, тогда как повышение температуры от
30 до 40 °С увеличивает интенсивность
дыхания менее значительно (Q10 около
1,5). Это может быть связано с тем, что повышение температуры ускоряет в большей степени ферментативные
процессы по сравнению с поступлением кислорода в клетки. В силу этого
возникает недостаток кислорода, что и
лимитирует процесс. В процессе эволюции
растения приспосабливаются к определенным температурным условиям. На характере
реагирования сказываются происхождение
растений, географический ареал их распространения.
Большое значение имеет
фаза развития растений. По данным Б. А. Рубина, на каждой фазе развития растений
для процесса дыхания наиболее благоприятны те температуры, на фоне которых обычно проходит эта
фаза. Изменение оптимальных температур при дыхании растений в зависимости от фазы их
развития связано с тем, что в процессе онтогенеза меняются пути дыхательного обмена. Между тем для
разных ферментных систем наиболее благоприятными являются разные температуры. Так,
температурный минимум работы цитохромов лежит выше по
сравнению с флавиновыми дегидрогеназами.
В этой связи интересно, что в более поздние фазы развития растений наблюдаются случаи,
когда флавиновые дегидрогеназы
выступают в роли конечных оксидаз, передавая водород
непосредственно кислороду воздуха.
Снабжение
кислородом. Кислород необходим для протекания дыхания — именно кислород является конечным акцептором электронов, движущихся по дыхательной цепи. Увеличение
содержания кислорода до 5—8% сопровождается повышением
интенсивности дыхания. Дальнейшее возрастание
его концентрации обычно уже не сказывается на интенсивности дыхания.
Однако из этого общего положения имеются
исключения. Снабжение растительных тканей и клеток кислородом зависит не только
от его содержания во внешней среде, но
и от скорости его поступления. Между тем часто проникновение кислорода к тем или иным тканям
затруднено. Это обстоятельство может проявляться на семенах и на плодах
с плотной оболочкой. В этом случае увеличение
концентрации кислорода в среде до
20% и более повышает интенсивность дыхания. Если семя гороха лишить
оболочки, то интенсивность дыхания возрастает с повышением содержания кислорода
в среде примерно до 5—10%. Однако
дыхание неповрежденных семян возрастает при увеличении содержания
кислорода до 20% и более.
Большое значение в снабжении кислородом
отдельных органов и тканей имеет система межклетников, способствующая
циркуляции воздуха. Воздух, проникая через устьица листа, достигает по межклетному пространству других органов, что и
позволяет им осуществлять аэробное дыхание. Доступ кислорода по межклетникам важен
для корневых систем растений, произрастающих на плохо аэрируемых почвах.
Известно, что приспособление корневых систем к росту в анаэробных условиях связано с развитием особенно большого объема межклетников. Вместе с тем нельзя
забывать, что корни многих растений не
имеют подобных приспособлений и для них очень важна хорошая
аэрация почвы. В отсутствие кислорода дыхание уступает место брожению. При содержании кислорода
ниже 5% брожение усиливается и выделение углекислого
газа начинает превышать поглощение
кислорода. Это приводит к тому, что
дыхательный коэффициент, как правило, становится больше единицы. При
повышении содержания кислорода процесс брожения полностью ингибируется (эффект Пастера) и дыхательный коэффициент
становится равным единице. Так, в опытах с плодами яблони было показано, что при снижении концентрации О2 выделение СО2 начинает расти.
Это увеличение выделения СО2 по сравнению с
поглощением О2 связано с усилением гликолиза и сопровождаемым спиртовым брожением. Вместе с тем добавление О2 ингибирует гликолиз. Необходимо отметить также, что кислород оказывает стимулирующее влияние на процесс фотодыхания.
Содержание
углекислого газа. СО2 является конечным
продуктом
как брожения, так и аэробного дыхания. При довольно высоких концентрациях СО2, значительно превышающих те, которые обычно окружают растительный организм (выше 40%),
процесс дыхания тормозится. Торможение вызывается несколькими причинами: 1) высокая концентрация СО2
может оказывать общее анестезирующее влияние на растительный организм;
2) СО2 тормозит активность ряда дыхательных ферментов; 3) повышение
содержания СО2 вызывает закрытие
устьиц, что затрудняет доступ кислорода и косвенно тормозит процесс дыхания.
Содержание воды. Небольшой водный
дефицит растущих тканей увеличивает интенсивность дыхания. Это связано с тем,
что водный дефицит и даже подвядание листьев
усиливают процесса распада сложных углеводов (крахмала) на более простые (сахара). Увеличение
содержания Сахаров этого основного субстрата дыхания усиливает сам процесс. Вместе с
тем при водном дефицит нарушается сопряжение окисления и фосфорилирования. Дыхание в этих условиях представляет в
основном бесполезную трату сухого вещества. При длительном завядании
растение расходует сахара интенсивность
дыхания падает. Иная закономерность
характерна для органов, находящихся в состоянии покоя. Увеличение содержания воды в семенах с 12 до 18% уже
увеличивает интенсивности дыхания в 4 раза. Дальнейшее повышение содержания
воды до 33 % приводит к увеличению интенсивности дыхания примерно в 100 раз. При перемещении растения или ткани из воды в
раствор солей дыхание усиливается — это так называемое солевое
дыхание.
Свет. Вопрос о влиянии света
на интенсивность дыхания изучался многими физиологами. Решение этого вопроса осложнено
методическими
трудностями. На свету трудно отделить процесс фотосинтеза от процесса дыхания.
Трудно разграничить прямое и косвенное влияние света. Так, на свету идет
фотосинтез. В свою очередь, влияние фотосинтеза на дыхание может быть различным
и даже
противоположным. Так, в процессе фотосинтеза образуются основные субстраты дыхания —
углеводы. Вместе с тем промежуточные
продукты, образовавшиеся при дыхании,
могут вовлекаться в фотосинтетический
цикл. Установлено, что свет стимулирует особый процесс фотодыхания. Все же применение метода меченых атомов позволило, хотя и не полностью, отграничить
процесс фотосинтеза от дыхания. В
настоящее время полагают, что влияние света
на процесс дыхания многообразно. Под влиянием света, особенно коротковолновых сине-фиолетовых лучей,
интенсивность обычного темнового дыхания возрастает. Активация дыхания светом показана на бесхлорофилльных растениях. Возможно
также, что свет активирует дыхательные
ферменты (оксидазы).
Питательные
соли. Интенсивность
дыхания сильно зависит от снабжения растения элементами минерального питания. Такие
элементы, как фосфор, сера,
железо, медь, марганец, принимают непосредственное
участие в процессе дыхания, входя в промежуточные продукты (фосфор) или
являясь составной частью дыхательных ферментов.
Поранение.
Поранение
органов и тканей растения усиливает интенсивность дыхания. Это связано с
разрушением клеток, из-за чего
повышается соприкосновение дыхательных субстратов и ферментов. Частично поранение может вызывать переход клеток в меристематическую фазу роста. Интенсивность дыхания делящихся клеток всегда выше по сравнению с закончившими рост.
2. ВЛИЯНИЕ ВНУТРЕННИХ ФАКТОРОВ НА ПРОЦЕСС ДЫХАНИЯ
Различные виды и
экологические формы растений дышат с разной интенсивностью. Как уже упоминалось,
светолюбивые растения характеризуются более высокой интенсивностью дыхания по
сравнению с теневыносливыми. Растения северных широт
дышат более интенсивно по сравнению с южными, особенно при
пониженной температуре. Интенсивность дыхания зависит от возраста. Как правило, более молодые растущие
органы и ткани дышат более интенсивно. Интенсивность дыхания проростков обычно
резко возрастает в течение
периода их наибольшего роста (первые 4—5 суток после начала прорастания), а затем начинает падать. По-видимому, это связано с образованием закончивших рост
тканей. Определенным закономерным
образом изменяется интенсивность дыхания листьев. После появления листа в первые дни его роста интенсивность дыхания возрастает, а затем резко падает,
а в период пожелтения часто вновь
немного повышается. Такое же явление наблюдается
у плодов перед их созреванием (климактерический период). Перед отмиранием организма или органа обычно наступает кратковременное усиление процесса дыхания.
Это связано, по-видимому, с
какими-то необратимыми процессами дегенерации тканей, при которых сложные соединения распадаются на более простые, что увеличивает количество субстратов
дыхания. В этот период дыхание не
сопровождается фосфорилированием. Коэффициент Р/О резко падает.
По-видимому, разрушается упорядоченное
расположение окислительных и фосфорилирующих систем.
Изменение возраста всего растительного
организма также сказывается на интенсивности дыхания. Наивысшей
интенсивностью дыхания обладают растения перед началом цветения. Органы растения, закончившие рост
или находящиеся в состоянии покоя, характеризуются низкой интенсивностью дыхания.
Очень низкое дыхание характерно
для сухих семян, закончивших рост плодов, тканей, в которых имеется большой процент мертвых клеток. Низкая интенсивность дыхания у покровных тканей. Высокой
интенсивностью дыхания характеризуются цветки (особенно тычинки и пестики),
клетки флоэмы, камбия. Как уже
упоминалось, различные органы и ткани растения сильно различаются по условиям
снабжения их кислородом. В листе
кислород свободно поступает практически к каждой клетке. Сочные плоды,
корнеплоды, клубни вентилируются очень
плохо; они слабо проницаемы для газов, не только для кислорода, но и для углекислого газа. Естественно, в
этих органах процесс дыхания
сдвигается в анаэробную сторону, дыхательный коэффициент возрастает.
Возрастание дыхательного коэффициента и сдвиг процесса дыхания в
анаэробную сторону наблюдаются в меристематических
тканях. Таким образом, разные органы характеризуются не только различной интенсивностью, но и неодинаковым качеством дыхательного процесса.
ПУТИ РЕГУЛЯЦИИ ДЫХАТЕЛЬНОГО ОБМЕНА
1.
ЛОКАЛИЗАЦИЯ В КЛЕТКЕ РЕАКЦИЙ ДЫХАТЕЛЬНОГО ОБМЕНА
Отдельные этапы
дыхания осуществляются в разных частях растительной клетки. Это определяется
распределением ферментов по отдельным органеллам с характерными для них
метаболическими функциями.
Изучение локализации, топографии ферментных систем имеет большое значение и для
понимания взаимодействия отдельных частей
клетки, а также возможности взаимодействия отдельных метаболитов. В
цитоплазме сосредоточены ферменты, катализирующие процесс гликолиза и
пентозофосфатного пути. Есть данные, что ферменты гликолиза имеются также в
матриксе митохондрий. Ферменты цикла Кребса
сосредоточены в основном в матриксе митохондрий. Ферменты дыхательной цепи
вплетены в определенной
последовательности во внутреннюю мембрану митохондрий. Приблизительно
20—25% общего белка во внутренней мембране
митохондрий составляют белки ферментов, участвующих в переносе водорода
и электронов. Предполагается, что ферменты-переносчики сгруппированы так, что каждая группа представляет самостоятельную
единицу — дыхательный ансамбль. В мембране митохондрий может быть
несколько тысяч таких ансамблей, которые равномерно распределены в мембранах.
Во внутренней мембране митохондрий локализованы также ферменты, обеспечивающие
процесс фосфорилирования
(АТФ-аза). Там же сосредоточен и переносчик
АТФ. Благодаря этому образовавшаяся в митохондриях АТФ может выходить из них и
использоваться в других частях клетки. Одновременно этот же переносчик осуществляет
перенос АДФ во внутреннее пространство
митохондрий. Через внутреннюю мембрану проникают
также пировиноградная кислота и некоторые органические кислоты цикла Кребса. Специфический переносчик осуществляет
перенос внутрь митохондрий фосфат-ионов. Вместе с тем
для коферментов НАД и НАДФ и некоторых
других веществ внутренняя мембрана
непроницаема.
Как рассматривалось выше,
часть никотинамидных коферментов восстанавливается
в цитоплазме в процессе гликолиза. Для того чтобы осуществить их окисление, существуют специальные механизмы.
У растений НАД-Н-дегидрогеназа, под
действием которой НАД-Н2 может
вступать в дыхательную цепь, локализована на наружной поверхности
внутренней мембраны. В случае отсутствия наружной НАД. Н-дегидрогеназы перенос НАД-Н2
через внутреннюю мембрану
осуществляется с помощью челночного механизма. Суть этого механизма следующая. Образовавшийся в цитоплазме НАД-Н2 реагирует с фосфодиоксиацетоном,
восстанавливая его до глицерофосфата.
Глицерофосфат проникает через мембрану и отдает водород флавиновой дегидрогеназе
и через нее в дыхательную цепь. При
этом глицерофосфат снова превращается в фосфодиоксиацетон, который выходит из митохондрии в цитоплазму и
опять подвергается восстановлению НАД-Н2. Сходный механизм
переноса энергетических эквивалентов через мембраны обнаружен и в хлоропластах. Таким образом, в клетке
одновременно осуществляется как разделение
веществ по разным компартментам, так и взаимосвязь между ними.
Важным является
вопрос, как обеспечиваются энергией процессы, происходящие в ядре клетки.
По-видимому, частично АТФ поступает
в ядро из цитоплазмы. В ядре имеются и собственные дыхательные ферменты. Так, в
нуклеоплазме обнаружены ферменты гликолиза. Есть
данные, что в ядре функционируют ферменты дыхательной
цепи, подобные митохондриальным. Наконец, ферменты дыхания обнаружены и в хлоропластах.
2. РЕГУЛЯЦИЯ ДЫХАТЕЛЬНОГО ОБМЕНА
Потребности организма
в энергии и метаболитах непрерывно меняются в зависимости от этапа его развития и
от условий среды, в которых он находится. В соответствии с этим на протяжении своего развития
организм способен изменять метаболизм, приспосабливаясь к изменяющимся условиям.
Вместе с тем в организме при всех условиях сохраняется взаимосвязанность всех
процессов обмена.
Все это достигается с помощью имеющихся в организме и клетке
специфических регуляторных механизмов. Подобные механизмы существуют и для регуляции путей
дыхательного обмена. Нерегулируемое дыхание
может привести к бесполезной трате сухого
вещества. Рассмотрим некоторые регуляторные механизмы. Как уже упоминалось, в присутствии кислорода
пировиноградная кислота претерпевает превращения по пути аэробного дыхания и перестает служить субстратом для процесса брожения
(эффект Пастера). Это связано с тем,
что для процесса брожения необходим НАД-Н2.
Между тем в аэробных условиях НАД-Н2 окисляется в дыхательной цепи и,
следовательно, не может быть использован на восстановление пирувата
до спирта. В результате под влиянием кислорода
процесс брожения затормаживается. В аэробных условиях уменьшается и скорость гликолиза, так как образуются значительные количества АТФ в процессе
окислительного фосфорилирования. Образовавшаяся АТФ тормозит активность фермента фосфофруктокиназы.
По-видимому, под влиянием АТФ меняется конфигурация
данного фермента. В связи с этим тормозится одна из первых реакций
гликолиза — превращения фруктозо-6-фосфата во фруктозе- 1,6-дифосфат. Эта регуляция имеет большой биологический смысл, так как ставит расход сахара в
зависимость от накопления АТФ. При
большем расходе АТФ ее содержание падает и гликолиз
ускоряется, при накоплении АТФ гликолиз тормозится. Отношение АДФ к АТФ имеет регуляторное значение и для реакций цикла Кребса.
Многие дегидрогеназы активируются под влиянием АДФ и одновременно ингибируются высокими
концентрациями АТФ. Здесь также
проявляется способность организма к регуляции своего энергетического
обмена. Важное значение имеет регулирование с помощью конечных продуктов реакций. Так, при
замедлении подачи активного ацетата (ацетил-КоА) в цикле Кребса щавелевоуксусная кислота накапливается. Это ингибирует активность фермента малатдегидрогеназы и тем самым приостанавливает работу цикла Кребса, предупреждая полное превращение всех интермедиатов цикла в ЩУК. Физиологи придают большое
значение регуляции соотношения распада глюкозы (гликолитический
или пентозофосфатный путь). Исходным
материалом как для того, так и для другого
пути служит глюкоза, поэтому в большинстве случаев между этими двумя
процессами имеются конкурентные взаимоотношения. Преимущественное осуществление того или иного пути связано с тем,
какой из ферментов — дегидрогеназа
глюкозо-6-фосфат или фосфогексокиназа —
образуется в первую очередь и в большем количестве.
Возможно, что этот тип регуляции осуществляется на уровне генома.
3. ВЗАИМОСВЯЗЬ ДЫХАНИЯ С ДРУГИМИ ПРОЦЕССАМИ ОБМЕНА
Процесс дыхания
представляет собой центральное звено обмена веществ организма, которое тесно связано с
другими процессами метаболизма. Необходимо подчеркнуть, что, несмотря на противоположную направленность двух центральных
процессов растительного организма — фотосинтеза и дыхания и на их рассредоточенность в разных органеллах клетки, между ними существует
тесная взаимосвязь. Прежде всего, для
протекания процесса дыхания нужны органические вещества (субстраты). Такими
субстратами в первую очередь являются углеводы, которые у зеленых растений образуются в процессе фотосинтеза. Вместе с
тем превращение веществ в процессе фотосинтеза
и дыхания идет через ряд сходных
промежуточных продуктов. Особенно много сходного в превращениях между фотосинтетическим циклом Кальвина и
реакциями пентозофосфатного пути
дыхательного обмена. Как в том, так и в другом случае происходят
взаимные превращения Сахаров с разной длиной углеродной цепочки (3, 4, 5, 6 и 7
углеродных атомов). По - видимому, несмотря на различное распределение (компартментацию) этих метаболитов в клетке, между ними
существует обмен. Иначе говоря,
промежуточные продукты дыхания могут быть использованы в процессе
фотосинтеза. Одновременно возможен и обратный
процесс. Много общего в энергетике фотосинтеза и дыхания в процессах
фотосинтетического и окислительного фосфорилирования.
Между этими двумя процессами возможен обмен энергетическими эквивалентами. АТФ, образовавшаяся на
свету при фотосинтетическом фосфорилировании, может служить основным источником энергии для различных биосинтетических
процессов, заменяя АТФ,
образовавшуюся в процессе дыхания. С другой стороны, АТФ и НАДФ-Н2,
образовавшиеся в процессе дыхания, могут быть использованы
для реакций цикла Кальвина. Имеются наблюдения, что на свету основными органеллами, поставляющими АТФ, являются хлоропласты.
Многие промежуточные
продукты процесса дыхания являются основой
биосинтеза важнейших соединений. Уже на протяжении первой, анаэробной фазы дыхания (гликолиз) триозофосфат, преобразуясь
в глицерин, может служить источником для синтеза жиров. Пировиноградная кислота путем аминирования
может дать аланин. Не менее важное значение
имеют и промежуточные продукты цикла
Кребса. Например, α -кетоглютаровая и щавелевоуксусная кислоты в процессе аминирования
дают аминокислоты —глутаминовую и аспарагиновую. Благодаря реакции переаминирования эти кислоты могут быть источником аминогруппы
для других аминокислот и, таким
образом, являться важнейшими промежуточными
продуктами для синтеза как белка, так и пуриновых и пиримидиновых
азотистых оснований. Янтарная кислота, образовавшаяся в цикле Кребса, дает основу для образования порфиринового ядра хлорофилла. Ацетил-КоА служит основой
для образования жирных кислот.
Поскольку имеется ряд реакций и процессов, благодаря которым отдельные компоненты извлекаются из цикла Кребса, должны быть и обратные процессы,
поставляющие их в цикл. Если бы этого не было, скорость превращения в аэробной фазе дыхания заметно бы снизилась. Такими
реакциями является окислительное дезаминирование
аминокислот, приводящее к образованию
органических кислот. Имеет значение также реакция карбоксилирования пировиноградной
кислоты или ее фосфорилированной формы, в результате
чего образуется щавелевоуксусная кислота.
Основной процесс, при котором образуются пентозы в растении,— это
пентозофосфатный путь дыхательного обмена. Пентозы входят в состав нуклеотидов, нуклеиновых кислот и ряда
коферментов, в том числе таких
важных, как никотинамидные (НАД и НАДФ), флавиновые (ФМН, ФАД). Пентозофосфатный путь дыхания
является также источником образования
эритрозо-4-фосфата. Эритрозофосфат, взаимодействуя с фосфоенолпируватом,
образует шикимовую
кислоту. Шикимовая кислота — материал для
образования ряда ароматических аминокислот, например триптофана, а из триптофана
образуется один из главных гормонов роста растений — ауксин (3-индолилуксусная кислота).
Рассмотренные связи дыхания и других
процессов метаболизма растения не являются
постоянными, раз навсегда данными. Они возникают
и нарушаются под влиянием как внутренних особенностей растения, так и внешних условий. При
неблагоприятных условиях эти
нарушения могут быть значительными и даже летальными.