4.2. Хромосома клеток эукариот

![]()
Схема строения хромосомы
в поздней профазе — метафазе митоза:
1—хроматида; 2—центромера; 3—короткое плечо; 4—длинное плечо.
Хромосо́мы (греч. χρώμα — цвет и греч. σώμα — тело) — хорошо
окрашиваемые включения в ядре
эукариотической клетки,
которые становятся легко заметными в определенных фазах клеточного
цикла (во время митоза или мейоза).
Хромосомы представляют собой высокую
степень конденсации хроматина, постоянно присутствующего в клеточном
ядре. Исходно термин был предложен для обозначения структур, выявляемых в
эукариотических клетках, но в последние десятилетия все чаще говорят о бактериальных
хромосомах.
Хромосомы
эукариот
Хромосомы эукариот имеют сложное строение. Основу хромосомы составляет
линейная (не замкнутая в кольцо) макромолекула дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, в молекулах ДНК хромосом
человека насчитывается от 50 до 245 миллионов пар азотистых оснований). В растянутом виде длина
хромосомы человека может достигать 5 см. Помимо нее, в состав хромосомы входят
пять специализированных белков
— H1, H2A, H2B, H3 и H4 (так называемые гистоны) и ряд негистоновых белков. Последовательность
аминокислот гистонов высококонсервативна и практически не различается в самых
разных группах организмов.
В интерфазе хроматин не конденсирован, но и в это время его
нити представляют собой комплекс из ДНК и белков. Макромолекула ДНК обвивает
октомеры (структуры, состоящую из восьми белковых глобул) гистоновых белков
H2A, H2B, H3 и H4, образуя структуры, названные нуклеосомами.
В целом вся конструкция несколько напоминает бусы. Последовательность из таких
нуклеосом, соединенных белком H1, называется нуклеофиламентом (nucleofilament), или нуклеосомной
нитью, диаметром около 10 нм.
В ранней интерфазе (фаза G1)
основу каждой из будущих хромосом составляет одна молекула ДНК. В фазе синтеза
(S) молекулы ДНК вступают в процесс репликации
и удваиваются. В поздней интерфазе (фаза G2) основа каждой из
хромосом состоит из двух идентичных молекул ДНК, образовавшихся в результате
репликации и соединенных между собой в районе центромерной последовательности.
Перед началом деления клеточного ядра
хромосома, представленная на этот момент цепочкой нуклеосом, начинает
спирализовываться, или упаковываться, образуя при помощи белка H1 более толстую
хроматиновую нить, или хроматиду, (chromatin
fiber) диаметром 30 нм. В результате дальнейшей спирализации диаметр хроматиды
достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр
1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой
микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными
плечами), поскольку две хроматиды, возникшие в результате репликации,
по-прежнему соединены между собой в районе центромеры (подробнее о судьбе
хромосом при клеточном делении см. статьи митоз и мейоз).
Центромера – первичная перетяжка
Вторичные
перетяжки
Морфологический признак, позволяющий
идентифицировать отдельные хромосомы в наборе. От первичной перетяжки
отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные
перетяжки бывают короткими и длинными и локализуются в разных точках по длине
хромосомы.
Типы строения хромосом
Различают четыре типа строения
хромосом:
·
телоцентрические (палочковидные хромосомы с центромерой, расположенной
на проксимальном конце);
·
акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным
вторым плечом);
·
субметацентрические (с плечами неравной длины, напоминающие по форме
букву L);
·
метацентрические (V-образные хромосомы, обладающие плечами равной
длины).
Тип хромосом является постоянным для
каждой гомологичной хромосомы и может быть постоянным у всех представителей
одного вида
или рода.
Спутники (сателлиты)
Сателлит - это округлое или удлиненное тельце
, отделенное от основной части хромосомы тонкой хроматиновой нитью, по диаметру
равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято
обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити
постоянны для каждой хромосомы.
Зона ядрышка
Зоны ядрышка (организаторы ядрышка) —
специальные участки, с которыми связано появление некоторых вторичных
перетяжек.
Хромонема
Хромонема — это спиральная
структура, которую удается увидеть в менее компактных хромосомах. Впервые
наблюдалась Баранецким в 1880 году в хромосомах клеток пыльников традесканции, термин ввёл Вейдовский. Хромонема может
состоять из двух, четырех и более нитей, в зависимости от исследуемого объекта.
Эти нити образуют спирали двух типов:
·
паранемическую (элементы спирали легко разъединить);
·
плектонемическую (нити плотно переплетаются).
Хромосомные
перестройки
Нарушение структуры хромосом
происходит в результате спонтанных или спровоцированных изменений.
·
Генные (точковые) мутации (изменения на молекулярном уровне);
·
Аберрации (микроскопические изменения, различимые при помощи
светового микроскопа):
o
делеции
o
инверсии
Гигантские хромосомы
Такие хромосомы, для которых
характерны огромные размеры, можно наблюдать в некоторых клетках на
определенных стадиях клеточного цикла. Например, они обнаруживаются в
клетках некоторых тканей личинок двукрылых
насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щеток). Именно
на препаратах гигантских хромосом удалось выявить признаки активности генов.
Политенные хромосомы
Впервые обнаружены Бальбиани в 1881 го, однако их
цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером.
Содержатся в клетках слюнных желез, кишечника, трахей, жирового
тела и мальпигиевых
сосудов личинок двукрылых.
Стандартная карта
политенной хромосомы разделяет геном на 102 пронумерованные полоски (1-20 -
X-хромосома, 21-60 - вторая хромосома, 61-100 - третья и 101-102 - четвёртая);
каждая из полосок поделена на 6 буквенных полосок (A-F), и каждая из
пронумерованных полосок также поделена на несколько (до 13) подразделов. На
фотографии выше указана 57-я полоска. Локализация многих генов известна точно
по буквенной шкале, но не точно по цифровой шкале, и поэтому указывается
приблизительно (например, 42C7-9, 60A1-2). Политенные полоски не имеют
одинаковой длины последовательности нуклеотидов, но в среднем одна буквенная
полоска содержит около 300kb ДНК, или 15-25 генов.
|
|
|
|
|
|
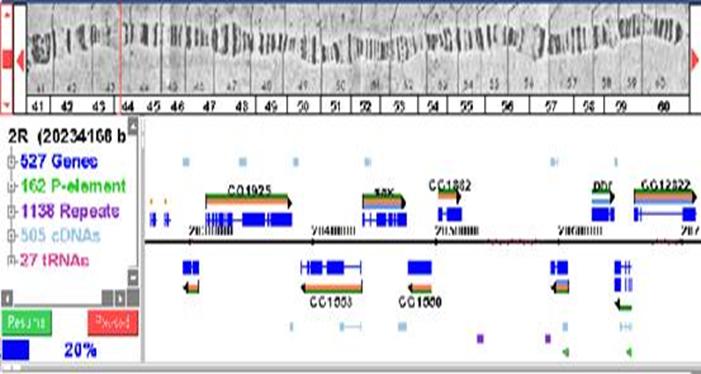
На рисунке - пример
компьютерной обработки (расшифровки) политенной хромосомы
Хромосомы типа ламповых щеток
Обнаружены Рюккертом в 1892 году. По длине
превышают политенные хромосомы, наблюдаются в ооцитах на стадии первого деления мейоза, во время которой процессы синтеза, приводящие к
образованию желтка,
наиболее интенсивны. Общая длина хромосомного набора в ооцитах некоторых
хвостатых амфибий достигает 5900 мкм.
Бактериальные хромосомы
Прокариоты (архебактерии и бактерии, в том числе митохондрии и пластиды,
постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого
слова. У большинства из них в клетке имеется только одна макромолекула ДНК,
замкнутая в кольцо (эта структура получила название нуклеоид). У ряда
бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо
нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме
прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так
называемых плазмид,
содержащих обычно незначительное, по сравнению с бактериальной хромосомой,
число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться
плазмидами в ходе парасексуального процесса.
Имеются данные о наличии у бактерий
белков, связанных с ДНК нуклеоида, но гистонов у них не обнаружено.